Frequently Asked Questions
|
FAQ Frequently Asked Questions |
What are the differences between Psykinematix v1.* (Standard & Student Editions) and v2.* (GPU & Bits # Editions)?
What are the limitations of the trial version?
What are the limitations of the demo or deactivated version?
Is Psykinematix compatible with the latest macOS?
Which version of macOS do you recommend using?
Is Psykinematix compatible with the Apple M Series processor found in Macintosh computers released since late 2020?
Can I run Psykinematix on my Windows-based PC?
Can I run Psykinematix on more than one computer?
What support is available?
I downloaded the Demo/Trial/Beta version but it does not work!
How accurate is Psykinematix timing?
What is the contrast resolution provided by Psykinematix?
What image, movie, and sound formats should I use?
How can I produce images or movies of my stimuli so I can include them in my papers or presentations?
Does Psykinematix include some image databases?
The experiments only run inside a window! How can I run them full-screen?
I'm sure that my display supports higher frame rates than those available in Psykinematix or in the Display System Preferences!
Does Psykinematix support multiple displays?
Which display calibration device is recommended?
Does Psykinematix support stereo modes?
Does Psykinematix support the Bits # device from Cambridge Research Systems?
Some of my input devices are not recognized! What's wrong?
Some unknown devices appear in the list! What's wrong?
My mouse has two buttons, but they are not recognized as left and right buttons! What's wrong?
How should I specify the space bar to terminate a stimulus presentation?
How should I disclose the use of Psykinematix in my publications?
Has anyone already made use of Psykinematix in published studies?
Version 1.9 of Psykinematix Student and Standard Editions, like all upgrades since 2008, is a free upgrade for all Psykinematix users and remains compatible with PowerPC-based Macintosh computers and Mac OS X 10.4 & 10.5, in addition to Intel-based Macintosh computers running Mac OS X 10.4 to macOS 10.14 for the 32-bit version and Mac OS X 10.7 to macOS 14 ("Sonoma") for the 64-bit version. These editions are also available as a Universal application to support both Intel 64-bit and M-Series Macintosh computers running macOS 11 ("Big Sur") to the latest macOS 14 ("Sonoma"). Although they do not support the most recent capabilities of modern graphics card, they are are the entry-level versions of Psykinematix for students and researchers on a budget and we will continue to provide regular & free updates for them.Version 2.* GPU Edition is a paid upgrade (Eligibility Checking Tool here) that brings support for real-time GPU-based stimulus generation and only works on Intel-based Macintosh computers running Mac OS X 10.6 or better, and M-Series Macintosh computers running macOS 11 or better (you should continue using v1.* otherwise). Version 2.* Bits#/Display++ Edition (also referred as 'Psykinematix #') is available exclusively from Cambridge Research Systems (CRS) and official resellers, and is dedicated to the Bits# and Display++ devices from CRS in addition to GPU support.
Note that upgrading your license to a new version (e.g. from v1.* to v2.*) still permits to run a previous version.
The main differences between the various Psykinematix Editions (Student, Standard, GPU, Bits#/Display++, Metropsis) are described in this comparison chart.
What are the limitations of the trial version?
The trial version is fully functional for 30 days. After this period, Psykinematix runs in Demo mode.
What are the limitations of the demo or deactivated version?
The demo and deactivated versions can still be used to create stimuli and experimental designs, but have the following limitations:Apart from these limitations, this demo version is fully functional making it the ideal FREE educational tool in classroom to introduce visual perception and illustrate psychophysical concepts to students.
- experiments can be run only in windowed mode (i.e. NOT fullscreen);
- video timing check and display calibration are not performed;
- stimuli preview saving is limited to 128 x 128 resolution;
- sessions cannot be exported as movies (when pressing the Control and clicking the Run button);
- session data are not saved into Psykinematix database.
Is Psykinematix compatible with the latest macOS?
Yes, all editions of Psykinematix are fully compatible with the latest macOS versions including the newly-released macOS 14 "Sonoma". See below for our macOS recommendation. We also provide both 32-bit and 64-bit versions of Psykinematix for Intel-based Macintosh computers as separate downloads. The 64-bit version is required for macOS 10.15 "Catalina" and above, and is still compatible with MacOS 10.7 to macOS 10.14. The 32-bit version is compatible with MacOS 10.4 to macOS 10.14 only. They are also available as Universal applications to support Intel 64-bit and M-Series Macintosh computers running macOS 11 ("Big Sur") to the latest macOS 14 ("Sonoma").
Psykinematix v1.* is compatible with Mac OS X 10.4 ("Tiger") or better, and runs on the older PowerPC-based (G4 and G5 CPUs), Intel-based, and M-Series Macintosh computers.
Psykinematix v2.* (GPU, Bits# & Metropsis Editions) is compatible with Mac OS 10.6 ("Snow Leopard") or better for Intel-based or M-Series Macintosh computers.
IMPORTANT NOTE: Before upgrading to a recent macOS version (10.7 or better) from an older version (10.6 or lower), make sure to de-activate your Psykinematix software using your license information. Then re-activate it once the new system has been successfully installed.
Which version of macOS do you recommend using?
In general, we do not recommend upgrading right away to the latest version of macOS because it may still be quite buggy, untested or not yet fully optimized for running demanding experiments.
Note that depending on the specific computer configuration and based on our in-house Psykinematix benchmark, newer versions of macOS may worsen OpenGL performance. If using an Intel-based Macintosh computer, we would recommend remaining on your current version of macOS if it already provides acceptable OpenGL performance for your specific computer configuration.
In short: Yes! See below for the long story:
Apple released its first Desktop processor, M1, specifically designed for the Mac in late 2020. Since then, the M2 and M3 processors have been released to power all Macintosh computers including the MacBook Air, MacBook Pro, Mac mini, iMac, Mac Pro and Mac Studio. Those Apple's M Series processors are, however, not compatible per se with softwares written to run on Intel processors found in earlier Macintosh computers.
To mitigate those software incompatibility issues, Apple has incorporated in its latest macOS versions (macOS 11 "Big Sur" or better) running on those new Macintosh computers a compatibility layer, called "Rosetta 2", that emulates Intel processors by translating Intel executable code into M Series executable code automatically and transparently to the user. Unfortunately this translation layer is not perfect and we found out that Psykinematix does not run properly under Rosetta 2 emulation. Rather than trying to fix these issues, we decided to provide native support for the M Series processor to fully take advantage of the new Apple silicon.
It took us longer than anticipated, but we have now fully native versions for most editions of Psykinematix that also run on the latest versions of macOS (Sonoma, Ventura and Monterey). They are tested daily with recent Macintosh computers equipped with the M1 and M2 processors (Mac mini M1, MacBookPro M1, MacBook Air M2 and Mac mini M2), and we will continue testing and supporting Psykinematix with the current M Series architecture (M3) and its future iterations.
Can I run Psykinematix on my Windows-based PC?
Psykinematix only runs on computers that support the macOS operating system from Apple; it does not run on Windows XP/Vista/7/8/10 operating systems from Microsoft. There is no plan to port Psykinematix to Windows or Linux because we think macOS is a better platform overall for running psychophysical experiments. Note that Macintosh computers can nowadays run either macOS, Windows or Linux which makes them the most versatile computers.
Can I run Psykinematix on more than one computer?
Yes, if you purchase a volume license or several single licenses but not if you only purchase a single license. The application is activated through the internet and is locked to the computer it was registered from, though it can be unlocked to transfer the registration to another computer. Note that it may be more advantageous for you to purchase a volume license rather than several single licenses (check the available license schemes on our online store).
Support is provided in priority to registered customers (i.e. those who purchased one or several licenses). High-priority and personalized support is also available through our Support Packages. Support may include troubleshooting, incident tickets, experimental designs, software and hardware customizations.
Non-registered users or those without support package can access the Psykinematix Forum or Psykinematix Facebook Group to get help from other users.
I downloaded the Demo/Trial/Beta version but it does not work!
Make sure your computer meets the minimum requirements for running Psykinematix (you may need to upgrade your system version to Mac OS X 10.4 or better) and to follow the installation and setup instructions. If your computer meets the minimum requirements, fill in a bug report by selecting "Feedback..." in the "Psykinematix" menu and detail precisely the problem you encountered and its circumstances. If you cannot use the software at all, contact us by e-mail at feedback@psykinematix.com.Psykinematix has successfully been tested on many Macintosh systems running Mac OS X 10.4 or better. Here is a non-exhaustive list (let us know if you had a successful experience on any other configuration):
Based on our in-house tests Psykinematix runs well on most MacPro, iMac, mac Mini, MacBookPro, MacBook and MacBook Air computers. However it is important to note that your computer configuration should match your experimental requirements in terms of video memory, graphics card power and CPU speed.iMac G4 FP 800Mhz
iBook G4 1Ghz
iMac G4 FP 1.25Ghz
PowerMac G4 1.25Ghz
MacMini G4 1.42Ghz
MacMini Intel Core 2 Duo 1.66 GHz
PowerBook G4 1.67Ghz
MacMini Intel Core 2 Duo 2 GHz
PowerMac G5 2.0GHz
iMac Core 2 Duo 2.16GHz
MacBook Pro Intel Core 2 Duo 2.33 GHz
iMac Intel Core 2 Duo 2.33 GHz
MacBook Pro Core 2 Duo 2.4 GHz
MacBook Pro Core 2 Duo 2.5 GHz
MacBook Pro dual-core Intel Core i5 2.3
Mac mini Intel Core i5 2.5 GHz
MacBook Pro Core i7 2.3 GHz
MacPro Quad-Core Intel Xeon 2x2.26 GHz
MacPro Quad-Core Intel Xeon 2x2.8 GHz
MacPro Quad-Core Intel Xeon 2x3.0 GHz
MacPro Quad-Core Intel Xeon 2x3.2 GHz
iMac 27" (late 2013) Quad-core Intel Core i7 3.5 GHz
iMac 27" Retina 5K (late 2014) Quad-core Intel Core i5 3.5GHz
Newest additions:
Mac mini (Late 2020), M1 8-Core 3.2 GHz
MacBookPro 13" (Late 2020), M1 8-Core 3.2 GHz
MacBookPro 14" (2021), M1 Pro 10-Core 3.2 GHz
Mac mini (Early 2023), M2 Pro 10-Core 3.5 GHz
How accurate is Psykinematix timing?
macOS is a multi-tasking operating system and there is the inherent risk of inaccurate timing. Timing accuracy depends on many factors, among them: processor and memory usage, operating system version, type of video card, graphics driver, network and other background activity, as well as the way Psykinematix manages timing. By default, Psykinematix runs at the highest priority during OpenGL operations and does its best to stay in sync with each display refresh (VBL or vertical blanking).Unreliable timing typically results in "missed/skipped" frames which may occur when Psykinematix competes with other programs in terms of CPU or memory usage, or for example when Psykinematix tries to draw too many textures into the OpenGL back buffer in preparation of the next frame presentation. However, Psykinematix carefully detects and reports the number of missed frames after each session as well as some statistics about the frame duration. See the Timing section for more details.
A fairly reliable experimental setup (with a frame rate of 60Hz) should produce near 0% of missed frames and no more than 0.1 ms for the standard deviation of the frame duration. This is assuming a minimal work-load (e.g. a single static stimulus). Increasing the frame work-load too much (i.e. with much more complex stimuli) will however eventually push your system to its limit and result in a significant number of missed frames.
What is the contrast resolution provided by Psykinematix?
Psykinematix supports by default the 8-bit per pixel frame buffer (256 levels per chromatic component) found in all macOS computers. For those with special needs (e.g. measurement of contrast threshold or presentation of sub-threshold stimuli), Psykinematix Standard & GPU Editions implement the following techniques to improve the luminance resolution (see Chromatic Modes in Defaults Preferences):
a bit-stealing technique that provides up to 11.6 bits of luminance resolution (Tyler 1997),
a spatial dithering technique that provides up to 14 bits of luminance and color resolution (Mulligan 1993),
a noisy-bit technique that provides a quasi-continuous resolution (Allard and Faubert 2008), up to 20 bits based on our study ("Influence of display type and rendering method on contrast sensitivity assessment").
The Bits#/Display++ Edition of Psykinematix ("Psykinematix Sharp") which supports the Bits# digital video processor and Display++ LCD monitor from Cambridge Research Systems (CRS) provide up to 14 bit and 16 bit monochromatic and chromatic outputs, respectively (see FAQ here).
What image, movie, and sound formats should I use?
All image, sound and movie formats recognized by QuickTime can be used with Psykinematix. If Psykinematix fails to open your media, try opening them with QuickTime. If QuickTime fails to open a file, Psykinematix will not be able to open it either. We recommend to use third-party software to convert your media to a QuickTime compatible format, like .mp4 for movie and .mp3 for sound.
The properties panel for most visual stimuli includes a preview image of your stimuli. Control-click the preview to open a contextual menu with various options, from which you can either:
copy the preview image (128 x 128 pixels by default) and directly paste it in the destination document,
save the stimulus to an image file in TIFF format at a size you specify,
save the stimulus as a movie in one of the available formats and at a size you specify.
In the absence of a preview, you may also run your experiment, and press and release the escape key (ESC) at the point of time when you like to capture a video frame, then select the “File → Save Screenshot As... → Image" menu to save the last captured frame as an image file in TIFF format. Use the "Preview" application to convert it to other image formats if necessary ("Preview" is part of Mac OS X and can be found in the “/Applications” folder).
You can also capture multiple consecutive video frames by holding the ESC key rather than releasing it immediately (the experiment presentation should then appear much slower), then select the “File → Save Screenshot As... → Movie" menu to save the captured frames as a movie file.
If you prefer to capture a whole session without having to press the ESC key, then press the CONTROL key while clicking the "Start" green button in the Designer panel. You will be then asked for the name and destination of the movie file.
Note however that in all cases, exporting a whole or partial experimental session will slow down the stimulus presentation and interfere with the video timing.
Does Psykinematix include some image databases?
No, Psykinematix does not include any image database you can directly use for your experiments. However here is a list of publicly available images databases you may find useful:Face Databases:
Natural Scenes Databases:
- The Japanese Female Facial Expression (JAFFE) Database: Lyons, Akamatsu, Kamachi & Gyoba (1998) Coding Facial Expressions with Gabor Wavelets. Proceedings, Third IEEE International Conference on Automatic Face and Gesture Recognition, April 14-16 1998, Nara Japan, IEEE Computer Society, pp. 200-205.
- The Karolinska Directed Emotional Faces (KDEF): Lundqvist, Flykt & Ohman (1998) The Karolinska Directed Emotional Faces - KDEF, CD ROM from Department of Clinical Neuroscience, Psychology section, Karolinska Institutet, ISBN 91-630-7164-9.
- Chicago Face Database: Ma, Correll & Wittenbrink (2015) The Chicago Face Database: A Free Stimulus Set of Faces and Norming Data. Behavior Research Methods, 47, 1122-1135.
- Face Research Lab London Set: DeBruine & Jones (2017)
- 100,000 Faces Generated by AI: Generated Photos (2019)
3D Objects Databases:
- The Berkeley Segmentation Dataset and Benchmark: Martin, Fowlkes, Tal & Malik (2001) A Database of Human Segmented Natural Images and its Application to Evaluating Segmentation Algorithms and Measuring Ecological Statistics. Proc. 8th Int'l Conf. Computer Vision Vol. 2, 416-423.
- McGill Calibrated Colour Image Database: Olmos & Kingdom (2004) A biologically inspired algorithm for the recovery of shading and reflectance images. Perception 33:1463-1473.
- The Barcelona Calibrated Images Database: Párraga, Vazquez-Corral & Vanrell (2009) A new cone activation-based natural images dataset. Perception 36(Suppl):180.
- Natural Scene Statistics Database for Vision Science from Center for Perceptual Systems, University of Texas at Austin: Geisler & Perry (2011) Statistics for optimal point prediction in natural images. Journal of Vision 11(12):article 14.
- Okazaki Synthetic Texture Image (OSTI) database: Okazawa, Tajima & Komatsu (2015) Image statistics underlying natural texture selectivity of neurons in macaque V4. Proceedings of National Academy of Sciences, USA 112:E351-E360.
See also:
- Amsterdam Library of Object Images (ALOI): Geusebroek, Burghouts & Smeulders (2005) The Amsterdam library of object images. Int. J. Comput. Vision 61(1):103-112.
- Gibson's Feelies - Shapes for haptic experiments: Norman, Phillips, Holmin, Norman, Beers, Boswell, Cheeseman, Stethen & Ronning (2012) Solid shape discrimination from vision and haptics: natural objects (Capsicum annuum) and Gibson's 'feelies'. Exp Brain Res 222:321-332.
- A Large Dataset of Object Scans (PLY format): Choi, Zhou, Miller & Koltun (2016) A Large Dataset of Object Scans. arXiv:1602.02481.
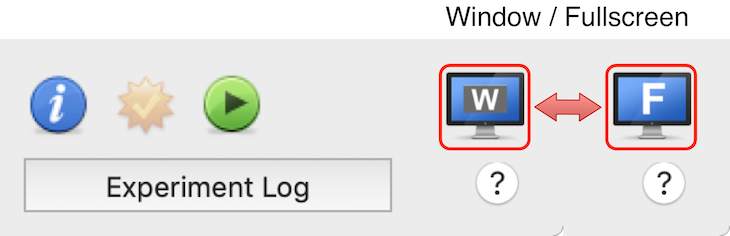
The experiments only run inside a window! How can I run them full-screen?
As illustrated below, there is a button in the bottom right corner of the Designer panel that allow to switch between the full-screen and windowed modes. Note that this button is grayed out if the demo or deactivated version of Psykinematix is running.
macOS provides by default a limited set of resolutions and refresh rates. There are third-party programs that allow you to adjust your Mac resolution and refresh rate to optimally match your monitor abilities:
SwitchResX (https://www.madrau.com/html/SRX/About.html)
- DisplayConfigX (https://www.3dexpress.de/)
Note that internal displays (LCD) in Macintosh laptops have typically a fixed frame rate of 60 Hz and most low-end external LCD monitors have a maximum frame rate of 75 Hz or 85 Hz. While high-end LCD or OLED monitors can nowadays reach frame rates as high as 120 Hz, 144 Hz or even higher, it is important to consider the timing issues discussed above when presenting complex stimuli.
Does Psykinematix support multiple displays?
Yes, but with one display at a time for stimulus presentation. Open the Display Preferences panel from Psykinematix' Preferences menu. All available displays and resolutions will be displayed. Select the one you wish to use as default for all your experiments. Each experiment can be later customized to run at at a specific resolution on a given display (Display tab from the Experiment properties).
Note that Psykinematix works very well with the Matrox DualHead2Go which provides an affordable solution by vertically splitting the stimulus screen into two parts, with each sent to a different display. This configuration can also be used for side-by-side stereo stimuli (Not supported in the Student Edition). Dual display presentation through multiple video cards may be supported in the future.
Which display calibration device is recommended?
Psykinematix has built-in support for several commercial colorimeters you can use for both luminance and color calibration (See the "Supported External Devices" chapter to learn more about the supported calibration devices).
If there is one device we would recommend, it is the affordable Spyder5 Express or SpyderX colorimeters from DataColor.
Does Psykinematix support stereo modes?
Yes, Psykinematix supports stereoscopic display based on either free viewing, mirror-based stereoscope, dual video outputs using the Matrox DualHead2Go video splitter (side by side or anamorphic format), anaglyph glasses, and "Frame Sequential Stereo" using for example a 3D graphics card such as the NVIDIA Quadro FX 4800 in combination with shutter glasses like REALD StereoGraphics CrystalEyes (see Stereo Rendering in Defaults Preferences).
Psykinematix also provides facilities to easily specify dichoptic or disparity-based stimuli (see the Visual Stimuli section).
The Bits#/Display++ Edition of Psykinematix (Psykinematix #), available exclusively from Cambridge Research Systems (CRS) and its official resellers (Namoto Trading in Japan), provides full support for the Bits# device and Display++ LCD monitor including CRS' infrared response boxes (CB6 and CT6) and ColorCal2 colorimeter. Note that support for the old Bits++ is now deprecated and will remain in "beta" stage indefinitely. We provide no assurance that Psykinematix v1.* or v2.* will work on your setup with the discontinued Bits++.
Some of my input devices are not recognized! What's wrong?
Psykinematix supports standard USB or Bluetooth mouse and keyboard devices that are HID-compliant as well as buttons present on Joystick or Gamepad USB HID-compliant devices.
Make sure that all your devices are correctly connected and functional before launching Psykinematix (some USB devices may need to be plugged into a powered USB hub). Disconnecting them while the application is running may cause instability problems. You should also check the list of available devices on the "Response Devices" panel of the Preferences window, and make sure the "Active" box of each of your input devices is checked. See the "Supported External Devices" chapter to learn more about the supported input devices.
Some unknown devices appear in the list! What's wrong?
Some unknown devices may appear in the list of available devices on the "Response Devices" panel of the Preferences window. Those are likely wireless devices that can be recognized through a USB receiver. Be sure to uncheck the "Active" box for each of these "phantom" devices. Some trial-and-error troubleshooting may be required to identify and disable these "phantom" devices.
My mouse has two buttons, but they are not recognized as left and right buttons! What's wrong?
The left button is the standard single button on Apple mice. To be recognized as the right button, the second button has to be configured as "Right-Click" using the software provided with your mouse. If this interferes with how you use the mouse because you configured the second button to perform other tasks, you should install USBOverdrive (https://www.usboverdrive.com) to customize the mouse settings to each application (you may have to uninstall the original software and driver provided by the mouse manufacturer).
How should I specify the space bar to terminate a stimulus presentation?
Use the special tag 'SB' in the text field. For more details about specifying inputs, see the Specifying Inputs section.
How should I disclose the use of Psykinematix in my publications?
State the name of the software, version and build number followed by the company and location in parentheses as illustrated in the following examples:"Stimuli were generated and presented using the Psykinematix software v1.9.5 Standard Edition Build 1283 (KyberVision Japan LLC, Sendai, Japan, psykinematix.com)"."Stimuli were generated and presented using the Psykinematix software v2.6 GPU Edition Build 1283 (KyberVision Japan LLC, Sendai, Japan, psykinematix.com)"."Stimuli were generated and presented using the Psykinematix-powered Metropsis software Research Edition v2.6 Build 1283 (Cambridge Research Systems Ltd, Rochester, UK, www.psychophysics.uk)".You may also cite the following paper if relevant to your study:
Has anyone already made use of Psykinematix in published studies?
Yes, here is below an non-exhaustive list of publications and theses that have reported using Psykinematix (let us know if you do using our publication report page: https://www.psykinematix.com//publications/):Preprints / in press
Levine AT, Yuen K, Billawa S, Bridge L, Clausen S, Cole E, Wang V, Canavan S, Spencer L, Campbell C, Morland AB, Codina C, Buckley D, Baseler HA (2020) Retinotopic remapping in the visual system of deaf adults. doi: 10.1101/2020.01.31.923342
Arias DJ, Saint-Amour D (2019) Synesthesia does not help to recover perceptual dominance following flash suppression. doi: 10.1101/2019.12.12.873992
Publications
Motz CT, Chesler KC, Allen RS, Bales KL, Mees LM, Feola AJ, Maa AY, Olson DE, Thule PM, Iuvone PM, Hendrick AM, Pardue MT (2020) Novel Detection and Restorative Levodopa Treatment for Preclinical Diabetic Retinopathy. Diabetes 69(7): 1518-1527. https://doi.org/10.2337/db19-0869
Georgiou M, Singh N, Kane T, Robson AG, Kalitzeos A, Hirji N, Webster AR, Dubra A, Carroll J, Michaelides M (2020) Photoreceptor Structure in GNAT2-Associated Achromatopsia. Investigative Ophthalmology & Visual Science, Vol. 61(40). doi:https://doi.org/10.1167/iovs.61.3.40
Huchzermeyer C, Fars J, Stöhr H, Kremers J (2020) Neue Techniken zur Quantifizierung des Farbsinns bei Störungen der Zapfenfunktion – Cambridge-Color-Test und photorezeptorspezifische zeitliche Kontrastempfindlichkeit bei Patient/Innen mit heterozygoten RP1L1- und RPGR- Mutationen. Der Ophthalmologe, https://doi.org/10.1007/s00347-020-01119-0
Sicard V, Simard A, Moore RD, Ellemberg D (2020) Practice effect associated with the serial administration of the switch task and its implications in the assessment of sports-related concussion. Journal of Clinical and Experimental Neuropsychology, Vol. 42(9):965–973
Augustinaite S, Kuhn B (2020) Ca2+ Activity of Sensory Activated and Suppressed Layer 6 Corticothalamic Neurons Reflects Behavioral State. Current Biology, Vol. 30(20) P3945-3960.E5
Gravelle MNK, Vandewouw MM, Young JM, Dunkley BT, Shroff MM, Taylor MJ (2020) More than meets the eye: Longitudinal visual system neurodevelopment in very preterm children and anophthalmia. NeuroImage: Clinical (2020), doi: https://doi.org/10.1016/j.nicl.2020.102373
Seider TR, Porges EC, Woods AJ, Cohen RA (2020) An fMRI study of age-associated changes in basic visual discrimination. Brain Imaging and Behavior, https://doi.org/10.1007/s11682-020-00301-x
Sicard V, Lortie JC, Moore RD, Ellemberg D (2020) Cognitive Testing and Exercise to Assess the Readiness to Return to Play After a Concussion. Translational Journal of the ACSM: Summer 2020 - Vol. 5(11):1–9
Mao Y, Min SH, Chen S, Gong L, Chen H, Hess RF, Zhou J (2020) Binocular Imbalance in Amblyopia Depends on Spatial Frequency in Binocular Combination. Invest. Ophthalmol. Vis. Sci. 2020;61(8):7. doi: https://doi.org/10.1167/iovs.61.8.7.
Qiu SX, Caldwell CL, You JY, Mendola JD (2020) Binocular rivalry from luminance and contrast. Vision Research, Vol. 175:41–50. https://doi.org/10.1016/j.visres.2020.06.006
Patterson Gentile C, Aguirre GK (2020) A neural correlate of visual discomfort from flicker. Journal of Vision, Vol.20, 11. doi:https://doi.org/10.1167/jov.20.7.11
Ryan DS, Sia RK, Eaddy JB, Logan LA, Familoni JO, Beydoun H, Rodgers SB, Rivers BA (2020) Preliminary evaluation of the VIsion PERformance (VIPER) simulator. Military Medical Research volume 7, Article number: 2
Georgiou M, Robson AG, Singh N, Pontikos N, Kane T, Hirji N, Ripamonti C, Rotsos T, Dubra A, Kalitzeos A, Webster AR, Carroll J, Michaelides M (2019) Deep Phenotyping of PDE6C-Associated Achromatopsia. Investigative Ophthalmology & Visual Science December 2019, Vol.60, 5112-5123. doi:https://doi.org/10.1167/iovs.19-27761
Cooper PR, Mendola JD (2019) Abnormal sensory eye dominance in stereoanomalous subjects. Journal of Vision, Vol.19, 14. doi:https://doi.org/10.1167/19.13.14
Costa MF, Gaddi CM (2019) A power law study of the edge influence on the perceived filling-in brightness magnitude. Psicologia: Reflexão e Crítica 32: 17. https://doi.org/10.1186/s41155-019-0130-7
Anssari N, Vosoughi R, Mullen KT, Mansouri B (2019) Selective Colour Vision Deficits in Multiple Sclerosis at Different Temporal Stages. Neuro-Ophthalmology, doi:10.1080/01658107.2019.1615960
Alnawmasi M, Chakraborty A, Dalton K, Quaid P, Dunkley B, Thompson B (2019) The effect of mild traumatic brain injury on the visual processing of global form and motion, Brain Injury 33(10), doi:10.1080/02699052.2019.1641842
Piotrowska B, Alexandra Willis A (2019) Beyond the global motion deficit hypothesis of developmental dyslexia: A cross-sectional study of visual, cognitive, and socio-economic factors influencing reading ability in children. Vision Research 159:48-60
Cohen D, Goddard E, and Mullen KT (2019) Reevaluating hMT+ and hV4 functional specialization for motion and static contrast using fMRI-guided repetitive transcranial magnetic stimulation. Journal of Vision 19(11), doi:10.1167/19.3.11
Tsai LT, Jang Y, Liao KM and Chen CC (2019) Legibility Assessment of Visual Word Form Symbols for Visual Tests. Scientific Reports, Vol. 9, Article number: 3338
Labhishetty V, Chakraborty A and Bobier W (2019) Is blur sensitivity altered in children with progressive myopia?. Vision Research 154:142-153
Himmelberg MM, Wade AR (2019) Eccentricity-dependent temporal contrast tuning in human visual cortex measured with fMRI. NeuroImage 184(1):462-474
Zhang J, Zhuang J, and Yu KM (2018) Posterior chamber phakic intraocular lens for the correction of high myopic anisometropic amblyopia in adults. International Journal of Ophthalmology 11(11):1870-1874
Shuffrey LC, Levinson L, Becerra A, Pak G, Sepulveda DM, Montgomery AK, Green HL, Froud K (2018) Visually Evoked Response Differences to Contrast and Motion in Children with Autism Spectrum Disorder. Brain Sciences 8(9), 160; doi:10.3390/brainsci8090160
Kumaran N, Ripamonti C, Kalitzeos A, Rubin GS, Bainbridge JWB, Michaelides M (2018) Severe Loss of Tritan Color Discrimination in RPE65 Associated Leber Congenital Amaurosis.Investigative Ophthalmology & Visual Science, Vol.59:85-93. doi:https://doi.org/10.1167/iovs.17-22905
Mikulskaya E, Martin FH (2018) Contrast sensitivity and motion discrimination in cannabis users. Psychopharmacology, 1-11, doi.org/10.1007/s00213-018-4944-2
Benjamin AV, Wailes-Newson K, Ma-Wyatt A, Baker DH and Wade AR (2018) The effect of locomotion on early visual contrast processing in humans. Journal of Neuroscience 1428-17
Welbourne LE, Morland AB, Wade AR (2018) Population receptive field (pRF) measurements of chromatic responses in human visual cortex using fMRI. NeuroImage 167(15):84-94
Hamm LM, Chen Z, Li J, Dai S, Black J, Yuan J, Yu M, Thompson B (2017) Contrast-balanced binocular treatment in children with deprivation amblyopia. Clin Exp Optom.
Zhang J,Yu KM (2017) Femtosecond laser corneal refractive surgery for the correction of high myopic anisometropic amblyopia in juveniles. Int J Ophthalmol 2017;10(11):1678-1685
Harris LR, Sakurai K, Beaudot WHA (2017) Tactile Flow Overrides Other Cues To Self Motion. Scientific Reports 7, Article number: 1059
Kim YJ, Tsai JJ, Ojemann J, Verghese P (2017) Attention to multiple objects facilitates their integration in prefrontal and parietal cortex. Journal of Neuroscience, 2370-16
Hamm L, Chen ZD, Li JR, Black J, Dai S, Yuan J, Yu MB, Thompson B (2017) Interocular suppression in children with deprivation amblyopia. Vision Research 133:112–120
Zhang J, Li JR, Chen ZD, Yu MB, Yu KM (2016) Phakic posterior chamber intraocular lens for unilateral high myopic amblyopia in Chinese pediatric patients. Int J Ophthalmol. 9(12):1790–1797
Bruce C. Hansen BC, Haun AM, Johnson AP, Ellemberg D (2016) On the Differentiation of Foveal and Peripheral Early Visual Evoked Potentials. Brain Topography 29(4):506–514
Chen Z, Li J, Liu J, Cai X, Yuan J, Deng D, Yu M (2016) Monocular perceptual learning of contrast detection facilitates binocular combination in adults with anisometropic amblyopia. Scientific Reports 6, Article number: 20187
Ding Z, Li J, Spiegel DP, Chen Z, Chan L, Luo G, Yuan J, Deng D, Yu M, Thompson B (2016) The effect of transcranial direct current stimulation on contrast sensitivity and visual evoked potential amplitude in adults with amblyopia. Scientific Reports 6, Article number: 19280
Tachibana R, Beaudot WHA, Sakurai K (2016) The range of discrepant directions indicated by visual and vestibular inputs that can be multimodally integrated in the perception of self-motion. Tohoku Psychologica Folia 75:9-14
Borges VM, Danesh-Meyer HV, Black JM, Thompson B (2015) Functional effects of unilateral open-angle glaucoma on the primary and extrastriate visual cortex. Journal of Vision, 15(9), doi:10.1167/15.15.9
Pageau M, de Guise D, Saint-Amour D (2015) Random-Dot Stereopsis in Microstrabismic Children: Stimulus Size Matters. Optometry & Vision Science 92(2):208-216
Chakraborty A, Anstice NS, Jacobs RJ, Paudel N, LaGasse LL, Lester BM, Wouldes tag, Harding JE, Thompson B (2015) Global motion perception is independent from contrast sensitivity for coherent motion direction discrimination and visual acuity in 4.5-year-old children. Vision Research 115 Part A:83-91
Tsai LT, Su YC, Beaudot WHA, Yang YH (2015) Assessment of Visual Dysfunction in a Patient with Right Occipital Infarct. Taiwan Journal of Physical Medicine and Rehabilitation 43(3):191-200
Li J, Spiegel D, Hess RF, Chen Z, Chan L, Deng D, Yu M, Thompson B (2015) Dichoptic training improves contrast sensitivity in adults with amblyopia. Vision Research
Baldwin AS, Husk JS, Edwards L, Hess RF (2015) The efficiency of second order orientation coherence detection. Vision Research 109A:45-51. doi:10.1016/j.visres.2015.01.026
Putnam CM, Bassi CJ (2015) Macular pigment spatial distribution effects on glare disability. Journal of Optometry
Chen Z, Li J, Thompson B, Deng D, Yuan J, Chan L, Hess RF, Yu M (2015) The Effect of Bangerter Filters on Binocular Function in Observers With Amblyopia. Investigative Ophthalmology & Visual Science 56:139-149. doi:10.1167/iovs.14-15224
Doualot A, Simard M, Saint-Amour D (2014) Attention modulation of stimulus rivalry under swapping paradigm. i-Perception 5(3):147-152. doi:10.1068/i0621
Zhou J, McNeill S, Babu RJ, Baker DH, Bobier WR, Hess RF (2014) Time Course of Dichoptic Masking in Normals and Suppression in Amblyopes. Investigative Ophthalmology & Visual Science 55:4098-4104. doi:10.1167/iovs.14-13969
LeMoyne E, Curnier D & Ellemberg D (2014) Pregnancy and cognition: Deficits in inhibition are unrelated to changes in fitness. Journal of Clinical and Experimental Neuropsychology. DOI:10.1080/13803395.2013.875520
Witz N, Zhou J & Hess RF (2014) Similar Mechanisms Underlie the Detection of Horizontal and Vertical Disparity Corrugations. PLoS ONE 9(1):e84846. doi:10.1371/journal.pone.0084846
Farrell-Whelan M & Brooks K (2013) Differential processing: Towards a unified model of direction and speed perception. Vision Research 92:10–18
Zhou J, Thompson B & Hess R (2013) A new form of rapid binocular plasticity in adult with amblyopia. Scientific Reports 3
Spiegel D, Byblow W, Hess R & Thompson B (2013) Anodal Transcranial Direct Current Stimulation Transiently Improves Contrast Sensitivity and Normalizes Visual Cortex Activation in Individuals With Amblyopia. Neurorehabilitation & Neural Repair 27(8):760-769
Witz N & Hess R (2013) Mechanisms underlying global stereopsis in fovea and periphery. Vision Research 87(19):10–21
Clavagnier S, Thompson B & Hess R (2013) Long Lasting Effects of Daily Theta Burst rTMS Sessions in the Human Amblyopic Cortex. Brain Stimulation
Peters M, Thompson B, Merabet L, Wu A & Shams L (2013) Anodal tDCS to V1 blocks visual perceptual learning consolidation. Neuropsychologia 51(7):1234–1239
Zhou J, Clavagnier S & Hess R (2012) Short-term monocular deprivation strengthens the patched eye's contribution to binocular combination. Journal of Vision 13(5):12
Huang PC, Li J, Deng D, Yu M & Hess R (2012) Temporal synchrony deficits in amblyopia. Investigative Ophthalmology & Visual Science
Huang PC, Baker DH, Hess RF (2012) Interocular suppression in normal and amblyopic vision: Spatio-temporal properties . Journal of Vision 12(11):29, 1–12
Li J, Thompson B, Ding Z, Chan LYL, Chen X, Yu M, Deng D & Hess RF (2012) Does partial occlusion promote normal binocular function? Investigative Ophthalmology & Visual Science 53
Husk JS, Huang PC & Hess RF (2012) Orientation coherence sensitivity. Journal of Vision 12(6):18, 1-15
Turgeon C, Champoux F, Lepore F, Ellemberg D (2012) Reduced visual discrimination in cochlear implant users. Neuroreport 23(6):385-389
Farrell-Whelan M, Wenderoth P, Brooks KR (2012) Challenging the distribution shift: Statically-induced direction illusion implicates differential processing of object-relative and non-object-relative motion. Vision Research 58:10-18
Johnson B, Wenderoth P (2011) Tapered dipoles in briefly flashed Glass-pattern sequences disambiguate perceived motion direction. Perception 40(4):383-391
Hess RF, Maehara G (2011) Does cognitive perception have access to brief temporal events? i-Perception 2(2):142-149
Hansen BC, Thompson B, Hess RF, Ellemberg D(2010) Extracting the internal representation of faces from human brain activity: An analogue to reverse correlation. NeuroImage 51(1):373-390
Dumoulin SO, Dakin SC, Hess RF (2008) Sparsely distributed contours dominate extra-striate responses to complex scenes. NeuroImage 42:890–901
Theses
Moreno Hernandez LM (2020) Visual awareness negativity and P3: ERP study of the distinctive neural correlates of conscious vision. Master Degree Program in Human Neuroscience, University of Turku, Finland
Lortie JC (2020) Les effets des impacts sous-commotions sur les fonctions exécutives et l'état psychoaffectif de jeunes hockeyeurs au cours d'une saison. Master - Maîtrise en sciences de l'activité physique, Université de Montréal, Québec, Canada
Coelho, Thais Fernandes Alvim (2019) Relação entre sensibilidade ao contraste e julgamento de expressões faciais de emoções em crianças de 6 a 14 anos [Relationship between contrast sensitivity and judgment of emotion facial expressions in children aged 6 to 14 years]. Master's Dissertation, Catálogo USP (São Paulo, Brazil)
Bakroon, Asmaa (2019) Psychophysical Studies Of Motion Perception In Autism Spectrum Disorders. PhD Thesis, University of Waterlo, Waterloo, Ontario, Canada
Zagui, Roberta Melissa Benetti (2019) Impacto da ambliopia estrabísmica e anisometrópica na visão de cores e de contraste espacial com diferentes níveis de complexidade (Impact of strabismic and anisometropic amblyopia on color and spatial contrast vision with different levels of complexity). PhD Thesis, University of São Paulo, Brazil
Levinson, Lisa Merideth (2018) Neural Correlates of Early-Stage Visual Processing Differences in Developmental Dyslexia. PhD Thesis, Columbia University, USA
Alnawmasi, Mohammed (2018) Investigating the Effect of Mild TBI on the Global Processing of Form and Motion. Master Degree in Vision Science, University of Waterlo, Waterlo, Canada
Labhishetty, Vivek (2017) Understanding the sensory and motor behavior of accommodation in progressive myopic children. PhD Thesis in Vision Science, University of Waterlo, Waterlo, Canada
Caldwell, Catherine L (2016) Perceptual Combination of Binocular Luminance and Orientation. Master Thesis, McGill University, Montreal, Canada
Welbourne, Lauren Elizabeth (2016) Peripheral factors affecting human colour perception. PhD Thesis, University of York, UK
Siong, Kar Ho (2016) Relationship between vision and balance in static and dynamic manners. PhD Thesis, The Hong Kong Polytechnic University, Hong Kong
Alamri, Sultan Ahmed H (2016) Neural Correlates of Gain Control in Drosophila and Humans. PhD Thesis, University of York, UK
LaRosa, Thomas J (2015) Correlational Evidence between the Processing Speed Index, Coherent Motion Threshold, and Achievement Scores of Children With and Without Learning Disabilities. PhD Thesis, University of Missouri, Saint Louis, USA
Chakraborty, Arijit (2015) Global Motion Perception in 4.5-year-old Children Born at Risk of Abnormal Neurodevelopment. PhD Thesis, The University of Auckland, New-Zealand
Putnam, Christopher (2014) Spatial Mapping of Macular Pigment Optical Density and Its Relationship to Contrast Sensitivity and Glare Disability. PhD Thesis, University of Missouri, Saint Louis, USA
Leung, Tsz Wing (2014) Characteristics of astigmatism in a Hong Kong Chinese population. PhD Thesis, The Hong Kong Polytechnic University, Hong Kong
Labonté-LeMoyne, Élise (2014) Effets de l'activité physique durant la grossesse sur le cerveau de la mère et de l'enfant. Thèse de Doctorat, Université de Montréal, Canada
Charbonneau, Yves (2014) L'impact de la commotion cérébrale d'origine sportive sur la capacité d'imagerie mentale visuelle d'athlètes. Thèse de Maîtrise, Université de Montréal, Canada
Gabbard, Stephen R (2013) Flash Lag Effect Model Discrimination. PhD Thesis, Wright State University, USA
Spiegel, Daniel (2013) Transcranial Direct Current Stimulation of the Healthy and Amblyopic Visual Cortex: Mechanisms and Action. PhD Thesis, The University of Auckland, New-Zealand
Witz, Nirel (2012) Properties of global stereopsis in fovea and periphery. Master Thesis, McGill University, Montreal, Canada
Kinerk, Wesley T (2012) Temporal Characteristics of Monoptic, Dichoptic and Half-Binocular Collinear Lateral Masking of Contrast Detection. PhD Thesis, University of Missouri, Saint Louis, USA
Turgeon, Christine (2011) Mesure du développement de la capacité de discrimination auditive et visuelle chez des personnes malentendantes porteuses d’un implant cochléaire. Thèse de Doctorat, Universitéa de Montréal, Canada
© 2006-2024 KyberVision Japan LLC. All rights reserved.
Mac, Macintosh, Mac OS, macOS and QuickTime are trademarks of Apple Inc., registered in the U.S. and other countries. Matrox, DualHead, and the names of Matrox products referenced herein are either trademarks and/or service marks or registered trademarks and/or service marks of Matrox. Cambridge Research Systems, Bits#, Bits++, and the names of Cambridge Research Systems products referenced herein are either trademarks and/or service marks or registered trademarks and/or service marks of Cambridge Research Systems. OpenGL is a registered trademark of Silicon Graphics, Inc. All other brand and product names are trademarks of their respective holders. Any omission of such trademarks from any product is regretted and is not intended as an infringement on such trademarks.